

До вашої уваги – ще один матеріал від авторів науково-популярного порталу «Моя наука» у нашій новій рубриці «Дивовижний світ науки». Портал створений для того, щоб з'являлося більше зрозумілих та якісних текстів українською мовою про наукові досягнення та відкриття, про українських дослідників. Тому й автори цієї рубрики – українські науковці у галузі біології та медицини.
Уявіть собі зиготу — клітину, яка утворюється при заплідненні, при злитті яйцеклітини та сперматозоїда. Із зиготи буде сформовано повністю весь зародок та частину плаценти у результаті великої кількості поділів клітин. Новий ембріон буде містити у собі велику кількість різноманітних тканин, які виконують найрізноманітніші функції, клітини яких не схожі одна на іншу: від нейронів до клітин шкіри та жирових клітин. І всі вони формуються від злиття яйцеклітини та сперматозоїда у зиготу. При чому переважну більшість маси зиготи формує саме жіноча статева клітина. Звідти можна було би припустити, що яйцеклітина неспеціалізована, що сама вона ніяка та не має жодних цікавих рис та характеристик — адже з неї після запліднення будуть сформовані всі можливі варіанти інших клітин. Але це припущення хибне: яйцеклітина має власну спеціалізацію і свої дуже цікаві особливості. Про деякі з них ми поговоримо сьогодні.
Яйцеклітина велика (набагато більша за сперматозоїд у своїх розмірах) та нерухома статева клітина. Але цікавим в ній є й те, що вона закладається ще в ембріональному періоді у ссавців. Тут слід додати, що не лише у ссавців є яйцеклітини та сперматозоїди — всі тварини, рослини та деякі гриби їх мають, у інших же організмів бувають однакові статеві клітини (ізогамія), й тоді їх не розділяють на яйцеклітини та сперматозоїди. Але у різних організмів відбувається різний процес дозрівання яйцеклітин. Так от, у ссавців оогенез — розвиток яйцеклітини (від лат. ovo — яйце) починається ще до народження дівчинки, перші стадії клітин-попередників яйцеклітини після народження і до підліткового періоду перебувають у стані спокою, а із статевим дозріванням починають по одній виходити зі стану спокою.
Але остаточне дозрівання яйцеклітини людини відбувається вже після запліднення. І, відповідно, якщо запліднення не відбулося, то така яйцеклітина не дозріває до фінальної стадії1.
Що це за стадії? У першу чергу, мова йде про зменшення кількості генетичної інформації, так званий мейоз. Справа в тому, що генетична інформація людини у будь-якій клітині представлена в подвійному наборі хромосом — 23 пари, разом 46 хромосом. Для того, щоб це число не подвоювалося з кожним наступним заплідненням, гамети — статеві клітини — повинні нести половину хромосом. Інакше дитина буде нести вдвічі більше хромосом за батьків, тобто 92, онуки — 184 тощо. Тому статеві клітини — це єдині клітини в організмі, які несуть половину генетичного матеріалу, тобто 23 хромосоми. І досягають вони цього завдяки мейозу, при цьому мейоз яйцеклітини та сперматозоїда відрізняється певними рисами.
Мейотичні поділи обидвох гамет, як чоловічої, так і жіночої, дають половинний набір хромосом, проте розміри клітини та час проходження поділів різні: з одного попередника утворюються однакові за розміром сперматозоїди і вони здатні утворюватися впродовж життя, тоді як мейоз яйцеклітин формує лише одну велику клітину з клітини-попередниці, й кількість цих клітин обмежена, а дозрівання відбувається раз на менструальний цикл.
Роздивимося детальніше мейоз яйцеклітини. Починається він так само, як і поділ інших клітин організму, наприклад, клітин шкіри — з подвоєння кількості хромосом (Рис. 1A). Це, на перший погляд, виглядає доволі парадоксально, — тільки що писали про те, що результатом мейозу є половинний набір хромосом — 23, — а тут пишемо, що підготовча стадія мейозу містить подвоєння їх кількості до 92. Але це просто означає, що потім поділів повинно бути не один — як у решті клітин при стандартному поділі, мітозі, (Рис. 1B) — а два, які дадуть чотири клітини з 23 хромосомами (Рис. 1C).
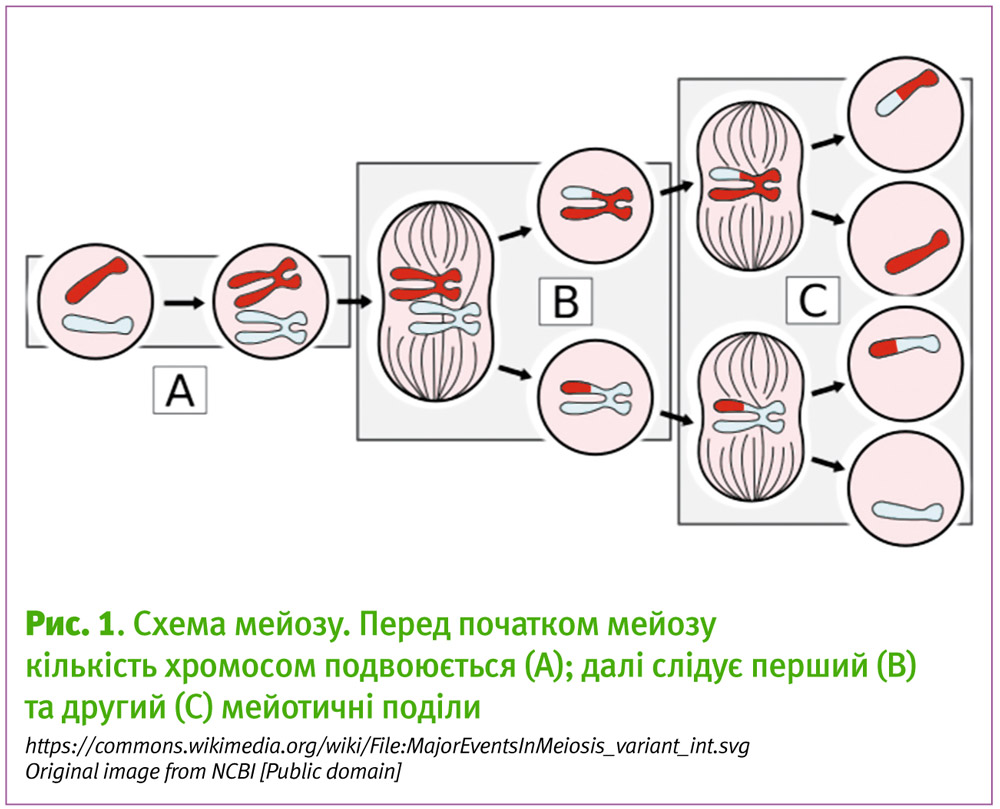
Причому у випадку мейозу жіночих статевих клітин, лише одна з них буде великою за розміром, бо поділ буде відбуватися таким чином, щоб вся маса цих клітин, весь вміст — цитоплазма — перейшла до одної дочірньої (Рис 2). Тоді як мейоз чоловічих статевих клітин дає чотири однакові за розміром сперматозоїда із 23 хромосомами в кожному2.
Отже, мейоз має 2 поділи. І тут у процесі дозрівання яйцеклітини можна говорити про фази: підготовка та початок мейозу відбувається до народження дівчини (Рис. 1 від A до B ліва частина), закінчується перший поділ мейозу при дії лютеінізуючого гормону після настання статевого дозрівання, тоді ж починається другий мейотичний поділ (Рис. 1 від B права частина, до C ліва частина), але при овуляції яйцеклітини не завершують його, натомість мейоз у людини повністю завершується після запліднення яйцеклітини.
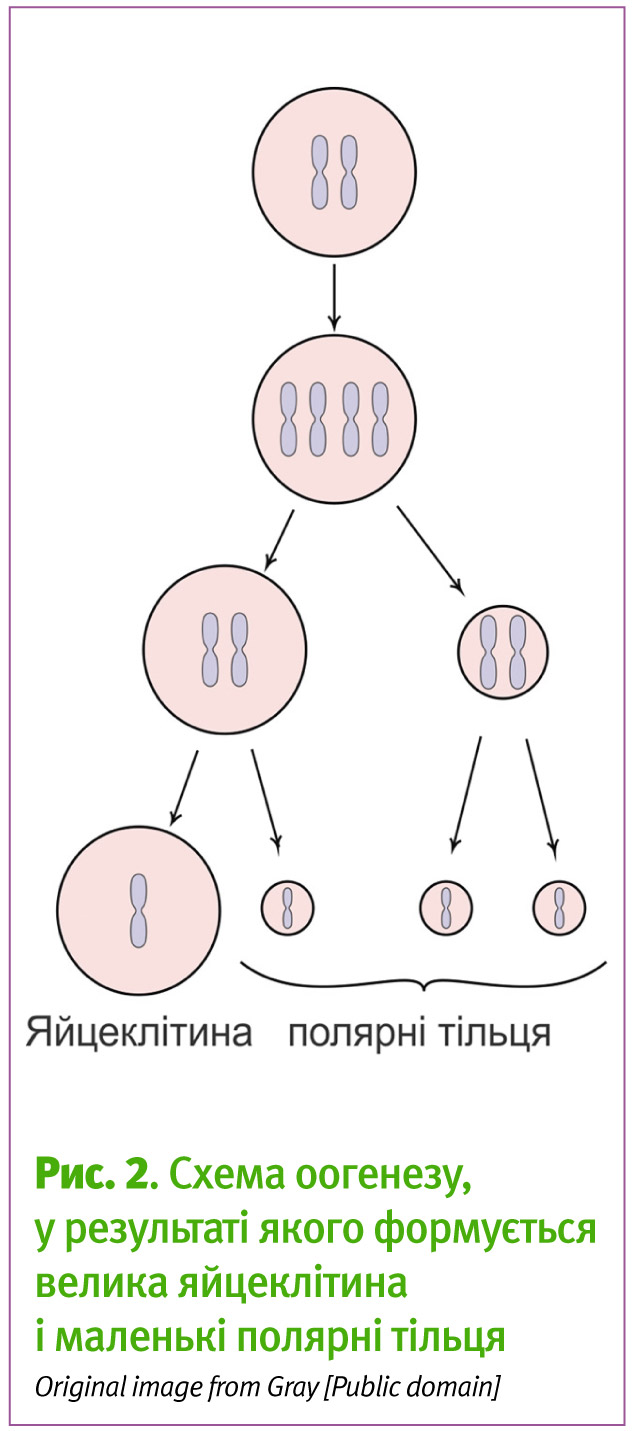
Але як це відбувається? Уявіть собі жіночу статеву клітину — вона куляста і доволі велика, у ній десь всередині містяться хромосоми, які є невеликими по відношенню до її загального об’єму. Їх треба розділити в два послідовних мейотичних поділи на 4 частини. При цьому основну масу цитоплазми потрібно залишити в одній з дочірніх, яка потім і стане власне яйцеклітиною, а іншим дочірнім клітинам, так званим полярним тільцям, роздати лише генетичний матеріал при поділах (Рис. 2 та 3)3. Такий розподіл дозволяє створити одну дуже велику клітину, а функції полярних тілець полягають лише у тому, щоб вивести додатковий генетичний матеріал і звести кількість хромосом яйцеклітини до 23.
Мейоз сперматозоїда у цьому питанні протікає легше, з топологічної точки зору — кожен раз слід поділити порівну на дві: як генетичний матеріал, так і цитоплазму, і це можна робити по центру клітини.
З яйцеклітиною так не вийде: кожен з поділів відбувається біля краю клітини, щоб одна з дочірніх клітин від’єднала лише частину цитоплазми разом з половиною хромосом і стала полярним тільцем (Рис. 3).
От ми й підійшли до центральної зірки поділу — центріолі.
Під час поділу — будь-якого поділу, як мітотичного, так і мейотичного — апарат клітини, який розтягує хромосоми навпіл до полюсів, де будуть формуватися дочірні клітини, називається веретеном поділу4. Таку назву він отримав тому, що за формою нагадує веретено — овальна форма з загостренням на протилежних кінцях, і тому, що містить в собі ниткоподібні формування — мікротрубочки. Згадані загострення на кінцях веретена поділу і є центросомами, основу яких складають центріолі. Від них тягнуться мікротрубочки у всі боки, до хромосом і до цитоскелету — основи каркасу клітини.
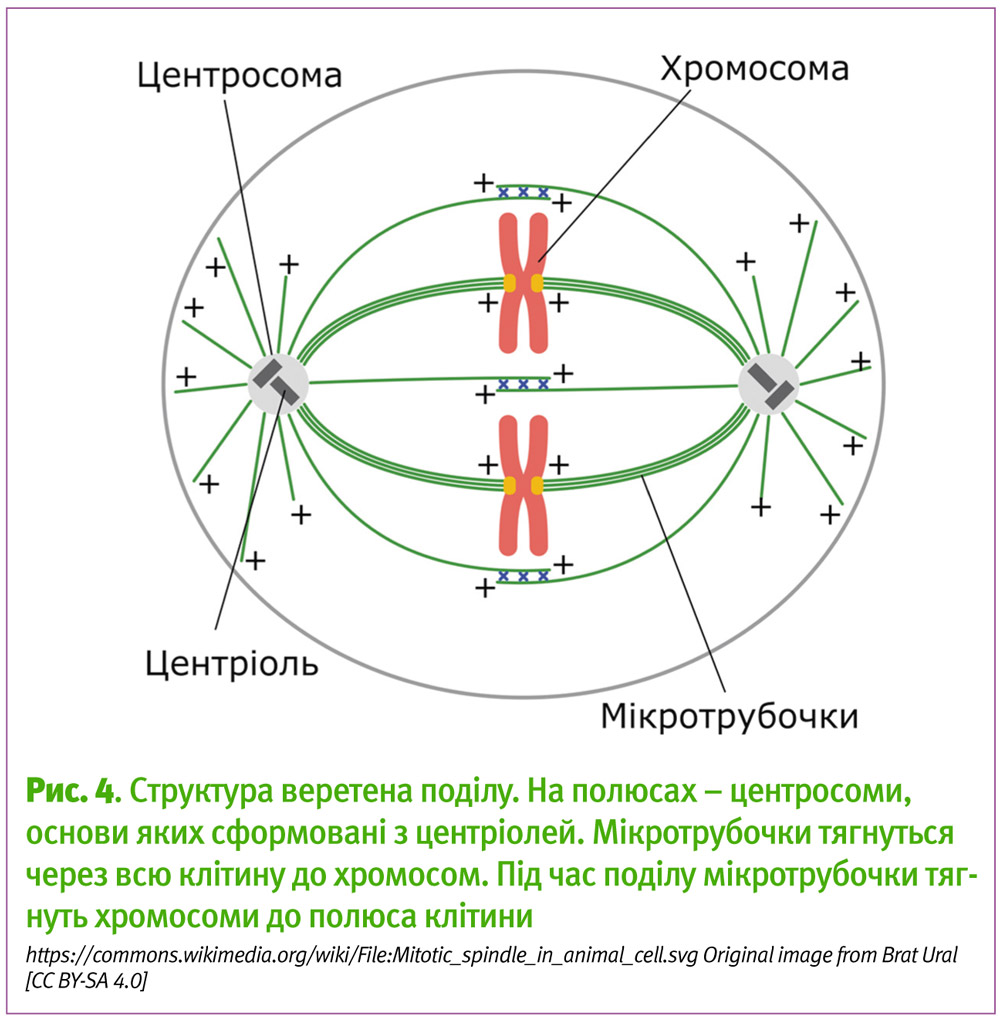
Під час поділу мікротрубочки тягнуть, як на буксирі, хромосоми до полюсів з центріолями, а сама структура веретена поділу щільно тримається за цитоскелет.
Таким чином, власне, і досягається розділення хромосом, у результаті якого половина хромосом скупчується навколо центросоми з одного боку, а інша половина — навколо протилежної центросоми.
Центріоль є базою для формування мікротрубочок, з неї вони ростуть у різні напрямки. Вони формують основу для цитоскелету клітини і є, так би мовити, «пупом» клітини. Їхня кількість доволі гарно контролюється, і в клітинах, що діляться, їх зазвичай дві — одна одразу після попереднього поділу, вона подвоюється і формує веретено поділу для наступного мітозу5. Також впорядкованість та налагодженість мікротрубочок необхідна для злиття генетичного матеріалу яйцеклітини та сперматозоїда після запліднення і формування нормальної зиготи.
Тому в процесі дозрівання яйцеклітин дуже цікавим і несподіваним є те, що центріолі деградують і відсутні під час мейозу. Натомість формування веретена поділу під час оогенезу відбувається без їхньої участі у не до кінця з’ясований спосіб — це так зване ацентросомне формування веретена поділу6.
Ця деградація може відбуватися різними шляхами. Так мухи Drosophila відділяють три центріолі у три полярні тільця під час мейозу, а центріоль яйцеклітини після завершення мейозу деградує. Проте оогенез у комах відбувається трохи по іншому від оогенезу ссавців (в тому числі й людини), а під час запліднення та завершення другого поділу мейозу у людини процес протікає вже без центріолей яйцеклітини. Стало відомо, що за відсутності центріолей основний центр збирання мікротрубочок веретена поділу – саме хроматин — білковий матеріал та ДНК хромосом7.
Взагалі, деградація центріолей є фундаментальною для яйцеклітин великого різноманіття організмів, від комах до людини, і згідно з основною теорією, вона запобігає самозаплідненню, або партеногенезу — формуванню зиготи шляхом злиття яйцеклітини і власного полярного тільця.
Порушення деградації центріолей під час оогенезу може призводити до порушень розподілення хромосом після запліднення. У нормі ж після запліднення центріолі у зародку формуються наново. При цьому велику частину матеріалу, з якого формується зародкова центріоль, приносить батьківський сперматозоїд.
На відміну від яйцеклітин, центріолі сперматозоїдів самців руйнуються не до кінця. При цьому основу рухового апарату хвоста сперматозоїда — базального тільця його джгутика – формує саме центріоль. Вона має трохи інший та додатковий білковий склад від центріолі, що входить до центросоми веретена поділу, проте саме сперматозоїд приносить основний сигнал для формування центріолі веретена поділу зиготи.
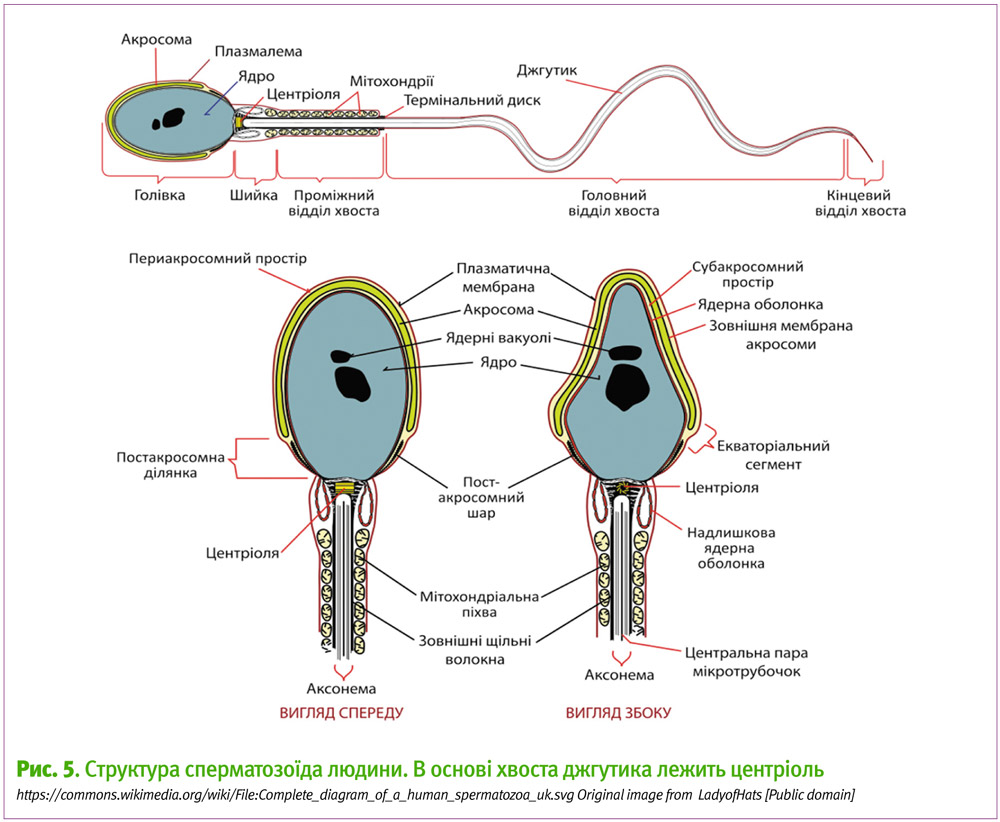
Так відбувається руйнування центріолей яйцеклітини, що становить великий теоретичний інтерес, проте потрібність компонентів центріолі сперматозоїдів має несподіваний та доволі цікавий практичний вихід у клініці. Адже під час штучного зовнішнього запліднення у пробірці сперматозоїди, які вколюють піпеткою до яйцеклітини, не проходять ті стадії, які вони зазвичай проходять при нормальному заплідненні, де протікає так звана акросомна реакція та вивільнення вмісту головки сперматозоїда у яйцеклітину, тоді як хвіст залишається ззовні. Натомість під час штучного запліднення клініцисти відрізають кінчик хвоста сперматозоїда для того, аби він не рухався, і вколюють його майже повністю, включаючи й основну частину хвоста, до яйцеклітини. Якби не відбувалося такої деградації центріолі яйцеклітини і включення компонентів батьківської центріолі вже в природі, то зовнішнє запліднення довелося б робити більш складними методами з руйнуванням основи джгутика, де міститься центріоль сперматозоїда. А так процес ін’єкції сперматозоїда відносно простий та не потребує сильних хімічних модифікацій сперматозоїда.
Але досі є невирішеною низка питань. Чи пошкодження формування основи джгутика сперматозоїда призводять до пошкоджених центріолей зиготи і відповідно можливих дефектів при мітотичних поділах зиготи — бластуляції? Чи при формуванні веретена поділу зиготи ці пошкодження можуть бути вилучені і покращені за допомогою білків, які знаходяться в цитоплазмі яйцеклітини? Хоча відомо, що для формування нової центріолі зародка потрібні певні компоненти центріолі сперматозоїда та додаткові компоненти з цитоплазми яйцеклітини, які діють разом і формують нормальну центросому для бластуляції та формування багатоклітинного зародка.
Але правильне формування веретена поділу та правильне розподілення хромосом під час формування гамет та після запліднення є дуже важливим для уникнення анеуплоїдій — неправильного набору кількості хромосом, що в більшості випадків залежно від хромосоми призводять до спонтанних абортів, або до трисомій, як то синдрому Дауна (трисомія по 21 хромосомі).
Також формування веретена поділу ооцита може проходити декількома різними механізмами, і залежить від організму. Так у людини й у миші ці шляхи відрізняються, тому при вивченні та доклінічних дослідженнях поламок формування веретена поділу варто використовувати такий модельний об’єкт, який має схожий до людського спосіб формування веретена поділу для дослідження людських патологій — те, що стало відомо лише у нещодавні роки8. Тому дослідження механізмів формування веретена поділу без центріолі в яйцеклітині потребує ще багато уваги від вчених у наступні роки.
Повний перелік літератури знаходиться у редакції.
- D. Clift and M. Schuh, “Restarting life: fertilization and the transition from meiosis to mitosis.” Nat. Rev. Mol. Cell Biol., vol. 14, no. 9, pp. 549–62, 2013.
- G. Manandhar, H. Schatten, and P. Sutovsky, “Centrosome reduction during gametogenesis and its significance.,” Biol. Reprod., vol. 72, no. 1, pp. 2–13, Jan. 2005.
- G. Coticchio and D. F. Albertini, Oogenesis. London: Springer London, 2013.
- S. L. Prosser and L. Pelletier, “Mitotic spindle assembly in animal cells: a fine balancing act” Nat. Rev. Mol. Cell Biol., vol. 18, no. 3, pp. 187–201, Feb. 2017.
- J. Loncarek and M. Bettencourt-Dias, “Building the right centriole for each cell type.” J. Cell Biol., pp. 1–13, Dec. 2017.
- P. T. Conduit, A. Wainman, and J. W. Raff, “Centrosome function and assembly in animal cells.” Nat. Rev. Mol. Cell Biol., vol. 16, no. 10, pp. 611–624, 2015.
- D. Inoue, J. Wittbrodt, and O. J. Gruss, “Loss and Rebirth of the Animal Microtubule Organizing Center : How Maternal Expression of Centrosomal Proteins Cooperates with the Sperm Centriole in Zygotic Centrosome Reformation,” vol. 1700135, pp. 1–9, 2018.
- S. Namgoong and N. Kim, “Meiotic spindle formation in mammalian oocytes: implications for human infertility †” vol. 98, no. May, pp. 153–161, 2018
коментарів