
До вашої уваги – ще один матеріал від авторів науково-популярного порталу «Моя наука» у нашій новій рубриці «Дивовижний світ науки». Портал створений для того, щоб з'являлося більше зрозумілих та якісних текстів українською мовою про наукові досягнення та відкриття, про українських дослідників. Тому й автори цієї рубрики – українські науковці у галузі біології та медицини.
Умови успішної посадки корабля Бластоциста на планеті Матка
Наталія Штефан,
портал «Моя наука» my.science.ua
У людини, порівняно з іншими видами, доволі низька фертильність. За даними досліджень1, максимальна ефективність зачаття на цикл становить приблизно 30%. Це означає, що серед 100 фертильних пар у період першого ж досліджуваного циклу відбувається 30 успішних зачать. Крім того, спостерігається доволі велика кількість випадків, коли вагітність виявляють за підвищеним рівнем хоріонічного гонадотропіну (гормон, який виділяється хоріоном ембріона після імплантації) та прогестерону жовтого тіла, але вона переривається до настання клінічно виражених проявів вагітності. Хоча при цьому може спостерігатись затримка менструального циклу всього на декілька днів або навіть не спостерігатись2. Це означає, що ембріон, швидше за все, ненадійно закріпився на внутрішній поверхні матки. Чому це так складно? Спробуємо розібратись.
І тут для полегшення сприйняття можна провести таку аналогію. Уявіть собі, що треба посадити корабель з екіпажем на поверхні планети. Зрозуміло, що для такої складної місії потрібно дуже багато. І одна невеличка поламка у будь-якій з ланок може бути причиною цілковитої невдачі. Важливо налаштувати всі «прилади», все добре підготувати, розрахувати запаси. Отже, що потрібно для успішного досягнення такої мети? По-перше, необхідно мати засоби зв’язку та контролю «посадки корабля». Такими засобами якраз і виступають: зі сторони матері – прогестерон, що виробляється жовтим тілом яєчника, та естроген (гормон фолікулярного апарату яєчників); зі сторони ембріону – хоріальний гонадотропін. Ці речовини глобально регулюють процес імплантації та подальшої вагітності. Цікаво, що вони впливають в першу чергу на матку. Отже, приймаюча «планета» також «знає», що до неї прибудуть гості. Їх потрібно влаштувати якнайкраще, бо вони гостюватимуть впродовж дев’яти місяців, і тому Матка ретельно готується до цієї події.
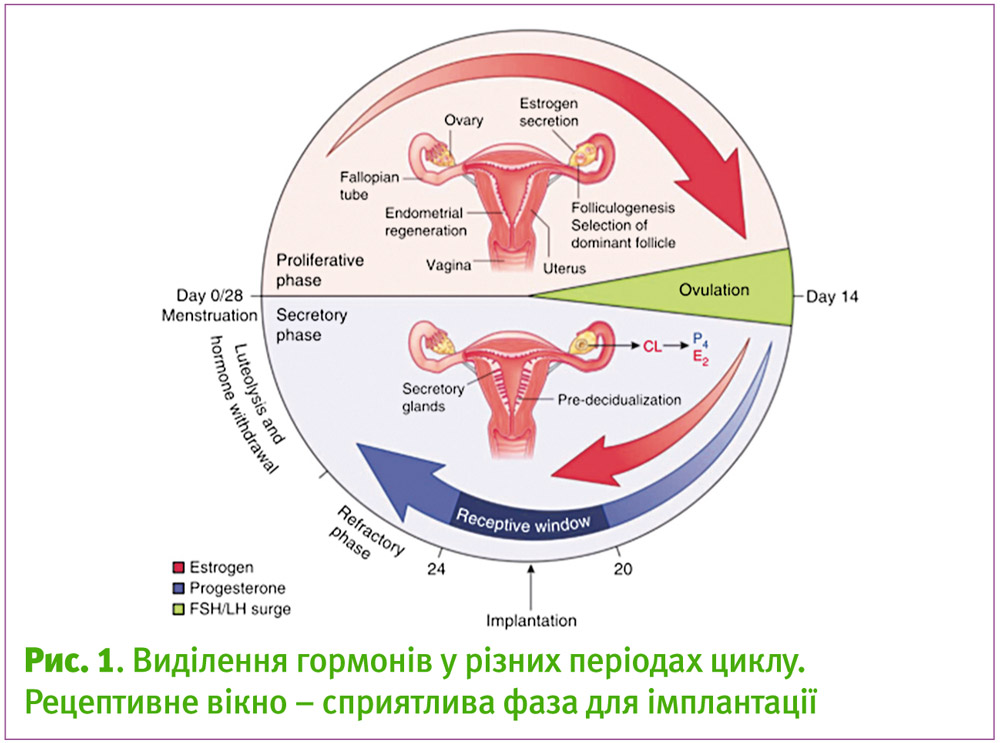
Під впливом різних гормонів впродовж менструального циклу відповідно існує три фази розвитку ендотелію: пререцептивна, рецептивна і нерецептивна (фаза відновлення). Ці фази були визначені як такі при дослідженнях на мишах, коли підсаджували ембріони. Відповідно, бластоциста може імплантуватись лише під час рецептивної фази, коли спостерігається формування мікровиступів на апікальній поверхні (піноподій) (рис. 2). У людини рецептивна фаза настає на 7 день після овуляції (що відповідає ранній лютеїновій фазі, при якій підвищується рівень прогестерону) і максимальна рецептивність досягається на середині лютеїнової фази (8–10 день після овуляції) (рис. 1). Далі у пізню лютеїнову фазу матка переходить у нерецептивну фазу аж до настання менструації. Зрозуміло, що першою структурою матки, яка контактує з бластоцистою, є епітелій порожнини матки, тому він відіграє основну роль у рецептивності та обміні сигналами між іншими частинами матки. Хоча деякі дослідження стверджують, що не менш важливу функцію відіграє строма, оскільки у ній відбувається значна зміна експресії генів. Вважають, що вона також може «керувати» рецептивністю епітелію3.
Експресія (генів) – процес, у ході якого спадкова інформація, закодована у вигляді ДНК, використовується для синтезу функціонального продукта – білка або РНК. Словом, якщо підвищується експресія якогось гену, то відбувається збільшення кількості білка, який закодований у послідовності цього гена. Так само зміна експресії може означати, що змінився певний набір білків у клітині, коли попередньо цей набір був іншим.
Оскільки за нашою аналогією з посадкою корабля гормони прогестерон та естроген є керувальними сигналами, то, відповідно, мають бути і прилади, що сприймають ці сигнали. Такими «приладами» є рецептори. Вони активуються у відповідь на зв’язування з гормонами та передають сигнал далі, що відповідно змінює їх функціональну активність. Активовані комплекси рецепторів прямують в ядро і взаємодіють там зі специфічними ділянками ДНК. Це, в свою чергу, призводить до активації або інгібування генів (регуляція експресії за допомогою білкових комплексів, що сприяють зчитуванню інформації з ДНК – транскрипційних факторів). Наприклад, до естрогену – це рецептори ERa та ERb. На мишах було доведено, що ERa критично важливий для дії естрогену, оскільки «вимкнення» гену цього рецептора призводило до повної нефертильності. Якщо ж вимикати гени рецепторів прогестерону, то виявляється, що рецептор PR-A є ключовим для імплантації, а при вимкненні PR-В спостерігається нормальна фертильність.
«Вимкнення» генів (генетичний нокаут) – генетична методика, при якій вибрані дослідником гени стають недієздатними або не функціональними (від терміну в боксі, коли боєць отримує настільки значний удар, що не може далі продовжувати змагання – нокаут).
Під впливом естрогену у проліферативній фазі відбувається ріст фолікулів, збільшується кількість клітин та строми епітелію, розвиваються залози епітелію. Рання секреторна фаза відзначається потовщенням ендометрію та формуванням жовтого тіла, яке згодом і виділяє прогестерон, що сприяє підготовці епітелію матки до імплантації – залози стають секреторними, спостерігається клітинна диференціація строми (предецидуалізація). У всіх досліджених видів прогестерон є абсолютно необхідним для імплантації. Без нього вона взагалі не відбувається. Однак, якщо подивитись на гормональний фон після овуляції (рис. 1), перед рецептивною фазою спостерігається ще й виділення естрогену. Високий рівень естрогену, що накладається на секрецію прогестерону, якраз і визначає рецептивне вікно (рис. 1) у середині лютеїнової фази на 20–24 дні циклу, що призводить до імплантації та подальшої вагітності. У мишей видалення яєчників перед тим, як відбулась преімплантаційна секреція естрогену, спричинює затримку імплантації та навіть призупинення розвитку ембріона (діапауза). Такий стан можна підтримувати впродовж декількох днів за допомогою штучного додавання прогестерону. А проведення ін’єкції естрогену може індукувати процес імплантації впродовж 24–48 годин після попереднього введення прогестерону. Це означає, що естроген запускає процеси імплантації на прогестероновому фоні.
Довгий час вважалось, що імплантація ембріона керується його власним призначенням і це пов’язано власне з його здатністю імплантуватися. Немає сумніву, що розвиток ембріона є критичним для досягнення успішної імплантації. Але найбільш свіжі докази свідчать про те, що ендометрій займає ключову позицію у керуванні часом та успішністю імплантації ембріона.
Після запліднення яйцеклітини відбувається декілька поділів. У порожнину матки ембріон потрапляє вже на стадії морули. Там він перебуває у «вільному плаванні» приблизно 2–3 дні перед тим, як прикріпитись до поверхні ендометрія.
Сама імплантація відбувається приблизно на 6–7 день запліднення, коли морула перейшла у стадію бластоцисти (рис. 2). мРНК материнського походження, яка обумовлювала синтез білків у період ранніх стадій ембріона, починає деградувати. Паралельно відбувається активація геному ембріона, інформація починає зчитуватись саме з генів ембріону.
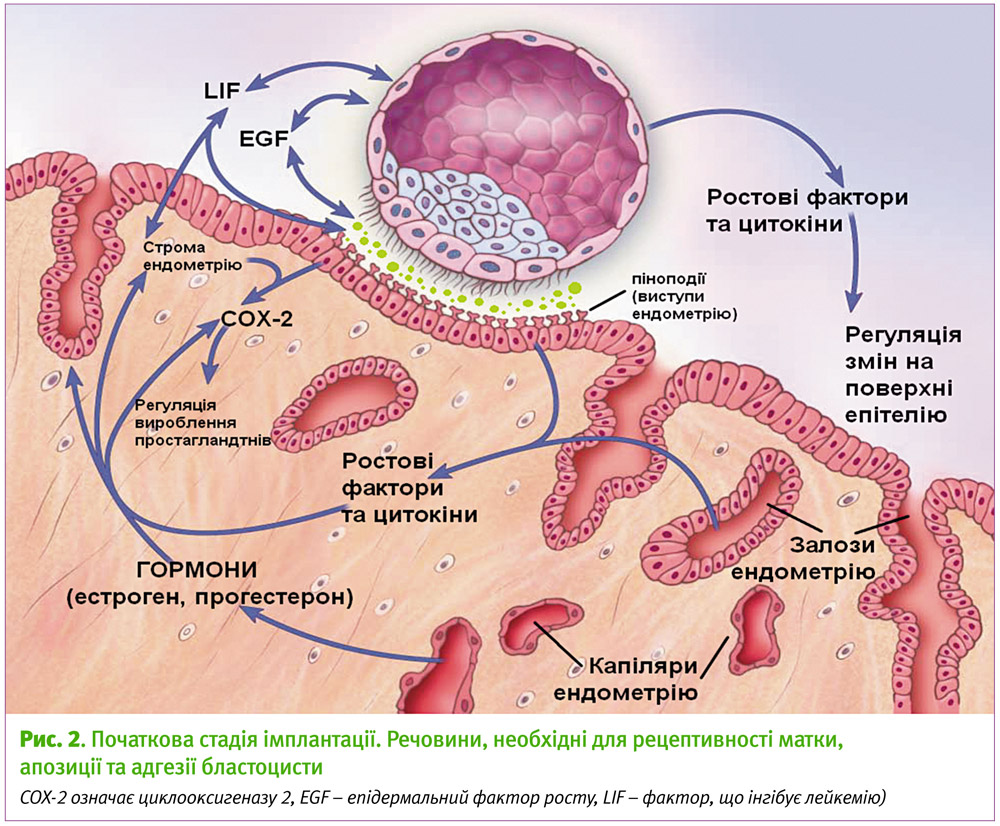
Перший фізичний контакт між готовою до імплантації бластоцистою та рецептивною маткою – апозиція – короткочасний, динамічний та нестабільний процес. На цьому етапі бластоцисті властиві ролінг (коли вона «котиться» по поверхні ендометрію) та хомінг (здатність клітин мігрувати у конкретні місця в організмі) в середину інвагінації, утвореної епітелієм. Мікроворсинки на апікальній поверхні трофобласту переплітаються із шароподібними виступами – піноподіями – на апікальній поверхні епітелію матки (рис. 2). До речі, формування піноподій корелює з максимальною рецептивністю ендометрія. Не дивлячись на те, що їхні функції залишаються нечітко з’ясованими, вважається, що вони функціонально важливі для переносу макромолекул, абсорбції рідин з порожнини матки та полегшення адгезії бластоцисти. Стабільна адгезія, на відміну від перших спроб нашого «корабля» зістикуватися з планетою, характеризується тісною фізичною взаємодією, між трофобластом та епітелієм формується функціональний взаємозв’язок, перебудовується цитоскелет клітин ендометрія, активуються різноманітні внутрішньоклітинні сигнальні каскади.
Важливу роль у процесі адгезії виконують інтегрини та кадгерини. Зокрема, інтегрини демонструють цикл-залежні зміни рівня експресії з підвищенням b3 та пониженням a4 субодиниць у період імплантаційного вікна. Наявність avb3 та avb5 інтегринів на апікальній поверхні епітелію є критичним для взаємодії між ендометрієм та трофобластом [4]. Інтегрини також присутні і на поверхні бластоцисти. Зокрема a5b1 субодиниця інтегрину під час активації бластоцисти переміщується на апікальну поверхню, що може бути важливим для початкового приєднання до епітелію матки4. Окрім цього, у процесі адгезії бере участь L-селектин, що знаходиться на поверхні бластоцисти. Він фізично з’єднується з молекулами (лігандами) на епітелії матки за принципом «ключ до замка». Усі взаємодії L-селектину, кадгеринів та інтегринів базуються на чіткій відповідності форми молекул, які мають об’єднатись і знаходяться на різних поверхнях.
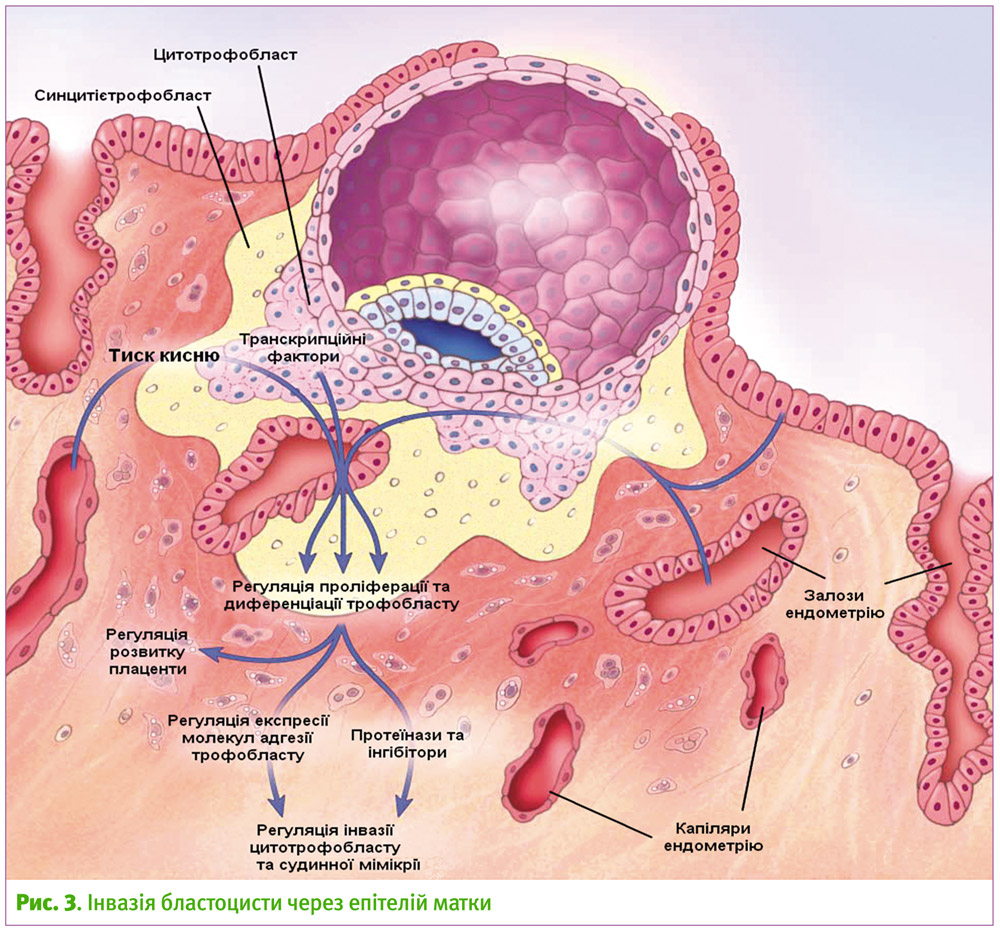
У багатьох ссавців приєднання бластоцисти збігається з підвищенням проникності судин в місці апозиції5 та децидуалізацією строми епітелію (поступове формування децидуальної оболонки). Епітеліальні клітини зміщуються від базальної мембрани, що дозволяє трофобласту поміщатись між клітинами епітелію, а згодом і проникати далі у строму (інвазія).
Крім того, нещодавні дослідження свідчать, що клітини трофобласту навіть здатні до ендоцитозу епітеліальних клітин (ентоз), що в буквальному сенсі означає поїдання ендометрію бластоцистою [4]. Закінчується інвазія повним вбудовуванням бластоцисти у строму ендометрія, а епітелій з боків відновлюється, щоб закрити місце імплантації (рис. 3). Під час інвазії клітини цитотрофобласту прагнуть дістатись спіральних артеріол матки. Це досягається завдяки тиску кисню, тому таким чином інвазія відбувається саме в артеріоли, а не венули [4]. Судини з високим опором та малими отворами в результаті стають судинами з великими отворами та низьким опором, що полегшує притік крові до ембріону. Вражаючою особливістю вторгнення цитотрофобласту є те, що він модулює експресію своїх молекул адгезії таким чином, щоб відповідати профілю ендотеліальних клітин, які йому зустрілись. Відбувається зменшення кількості рецепторів адгезії, характерних для стовбурових клітин цитотрофобласту (інтегрину a6b4, епітеліального кадгерину) та збільшення експресії рецепторів адгезії, характерних для ендотелію, включаючи кадгерини (судинний ендотеліальний кадгерин, кадгерин-11), рецептори родини імуноглобулінів G, а також інтегрини avb3 та a1b1. Такий процес називають «судинною мімікрією», що допомагає клітинам трофобласту уникати реакції материнської імунної системи.
Будь-який процес в організмі, пов’язаний із перебудовою або формуванням нового (а вагітність якраз і є таким процесом), неможливо уявити без факторів росту. У період імплантаційного вікна спостерігається їх підвищена експресія, особливо в місці апозиції ембріона (рис. 2). Серед них багато з родини епідермальних факторів росту (EGF). Доведено, що експресія рецепторів до EGF регулюється естрогеном та прогестероном. Один із таких рецепторів – НВ-EGF – стимулює ріст та розвиток бластоцист у лабораторних умовах (in vitro) та може індукувати експресію ендометріального інтегрину avb3, необхідного для приєднання бластоцисти.
Серед інших факторів росту одну з головних ролей у процесі імплантації відіграє LIF – фактор, що інгібує лейкемію (назва походить від його властивостей пригнічувати неконтрольоване розмноження лейкемічних клітин). Він належить до цитокінів (класу білкових молекул, які регулюють міжклітинні взаємодії в організмі, керують відповідями імунної та інших систем), родини IL-6 і контролює диференціацію клітин. Дія LIF, опосередкована через рецептори LIF-R та gp130, активує внутрішньоклітинний каскад реакцій, запускаючи функціональні відповіді, важливі для децидуалізації та імплантації. Активація LIF-R рецепторів у відповідь на зв’язування з LIF регулюється естрогеном та прогестероном. Естроген індукує секрецію LIF у залозах ендометрію. Вважається, що це основна мішень естрогену в ендометрії. Потрапляючи в порожнину матки, LIF зв’язується з рецепторами на поверхні епітеліальних клітин, що робить матку рецептивною для бластоцисти. Пік експресії LIF припадає на імплантаційне вікно, а у секреторній фазі не знайдено білків LIF. У мишей із вимкненими генами LIF спостерігалась нормальна овуляція, ооцити запліднювались, але ембріони не могли імплантуватись через дефективність процесу децидуалізації. Такий же ефект спостерігався при блокуванні гену рецепторів gp130 в матці. На противагу цьому, ембріони з вимкненими генами рецепторів LIF-R до LIF могли спокійно імплантуватись [4]. Це означає, що LIF може передавати сигнали іншими шляхами.
Відомо також, що у жінок із неплідністю та повторюваними викиднями спостерігається знижений рівень експресії у зразках рідин після промивання порожнини матки порівняно з фертильними жінками [4]. Окрім всього іншого, LIF приймає участь у диференціації цитотрофобласту6 у синцитієтрофобласт та сприяє секреції хоріонічного гонадотропіну.
Крім того, що для нашого «корабля» слід будувати щось нове під впливом факторів росту, йому потрібно розчистити «посадкове місце». У матці активно відбувається перебудова тканин, формування плаценти та оболонок плода, але для цього необхідно спочатку зруйнувати існуючі структури. Функцію руйнівників виконують спеціальні ферменти – матриксні металопротеїнази. Вони розщеплюють матрикс білків клітин ендометрію і присутні як в ендометрії, так і в клітинах ембріону. Основною металопротеїназою, яка регулює інвазію цитотрофобласту, є ММР-9.
Все вищеописане демонструє складність процесу імплантації. Хоча це далеко не все, що ми знаємо. І далеко не все, що ще можна дізнатись. Дивлячись на цю складну систему, легко уявити, як невеличкі поломки можуть призвести до небажаних наслідків. Нерідко трапляється, що імплантація не відбувається. Але якщо імплантація відбулась, але «корабель» сів криво, це буде відображатись впродовж усієї вагітності і в гіршому випадку призведе, знову ж таки, до викидня. Наприклад, у пацієнток із прееклампсією спостерігається неглибока інвазія цитотрофобласту і відповідно обмежена внутрішньосудинна інвазія. Такий цитотрофобласт не зміг перемкнути набір своїх молекул адгезії на такий, що схожий на набір у материнських ендотеліальних клітинах. І таким чином, замість того, щоб судини стали з великими отворами і низьким опором, вони залишаються такими, як були, з малими отворами та високим опором. Відповідно, у пацієнток спостерігаються підвищення тиску та клінічні симптоми прееклампсії.
Як бачимо, якщо щось пішло не так від самого початку, наслідки можуть бути глобальні. І не так вже й просто посадити корабель на планету на 9 місяців для життя та розвитку.
Повний перелік літератури
знаходиться у редакції.
- Zinaman MJ, Clegg ED, Brown CC, O’Connor J, Selevan SG. Estimates of human fertility and pregnancy loss. Fertil Steril [Internet]. 1996;65(3):503–9.
- Allen Wilcox, Clarice Weinberg, John O’Connor, Donna Baird, John Schlatterer, Robert Canfield, Glenn Armstrong BN. Incidence of early loss of pregnancy.
N Engl J Med. 1988;319. - Cha J, Sun X, Dey SK. Mechanisms of implantation: strategies for successful pregnancy. Nat Med [Internet]. 2012 Dec;18(12):1754–67.
- Farquharson RG, Stephenson MD. Early Pregnancy. 2017.
- Dey SK, Lim H, Das SK, Reese J, Paria BC, Daikoku T, et al. Molecular cues to implantation. Endocr Rev. 2004;25(3):341–73.
- Про структуру трофобласту і чому він такий, читайте у номері ЗТЖ №3(78) квітень 2017, «Віруси, які дарують нам життя»
коментариев