
Всім зрозуміло, що хворіти погано. І для власного організму та оточуючих (якщо це інфекція), і для майбутнього потомства. Але чи завжди це так? Та й взагалі, чи народились би ми, якби наші предки не хворіли інфекційними захворюваннями?
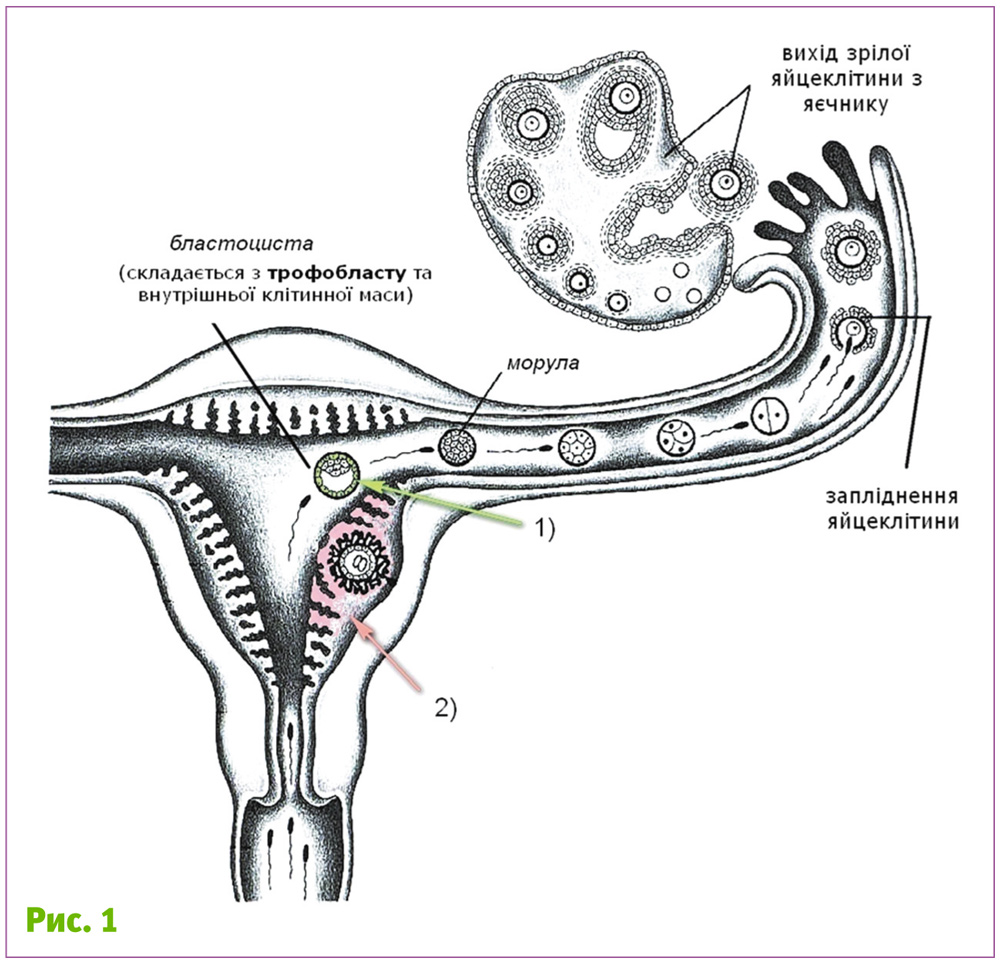
Щоб не розкривати карти одразу, підійдемо до цього питання здалека. Для початку згадаємо, як утворюється плацента. Цей тимчасовий орган, необхідний для живлення плоду через кров матері, утворюється із трофобласту (1 на рис. 1) та ендометрію (2 на рис. 1).
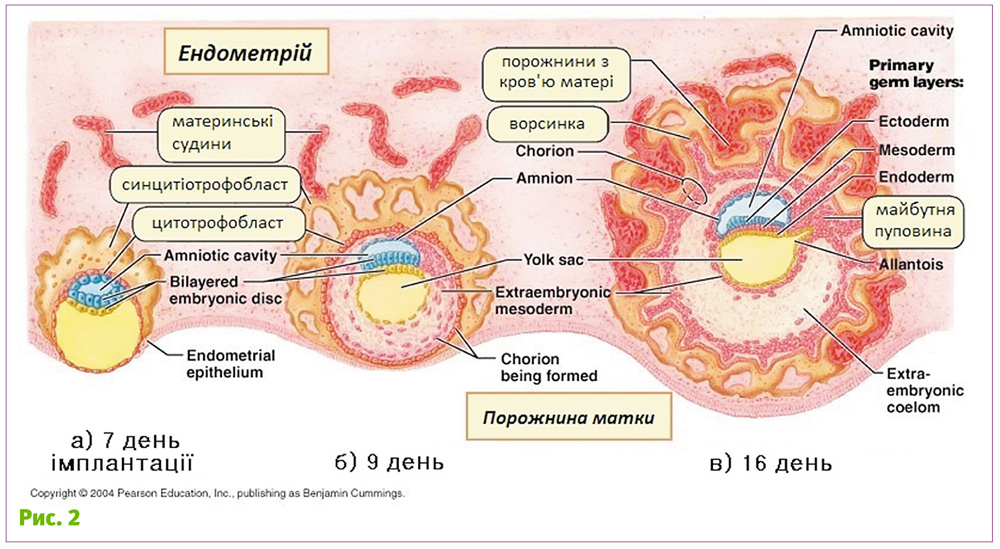
У людини та інших ссавців (але далеко не у всіх) під час імплантації ембріон вмуровується в слизову оболонку матки (рис. 2). При цьому деякі клітини трофобласту зливаються, утворюючи багатоядерну структуру – синцитій. З часом трофобласт приймає форму ворсинок, які поступово розгалужуються, руйнуючи слизову оболонку і судини матки у місці імплантації.
У подальшому ці ворсинки безпосередньо контактують з кров’ю матері. Отже, трофобласт (від грец. trophe – живлення, та blastos – зародок) – структура, через яку поживні речовини переходять від материнського організму до зародка. Найбільш близьким до матері виявляється саме синцитійний трофобласт, саме той зовнішній шар, де клітини злились. Саме ця структура безпосередньо контактує з організмом матері від початку імплантації аж до кінця вагітності. І зрозуміло, що від цього контакту залежить весь характер взаємодії «плід–мати».
Але чому клітини починають зливатись? Навіщо потрібен синцитій? Спробуємо знову підійти здалеку і згадаємо про віруси.
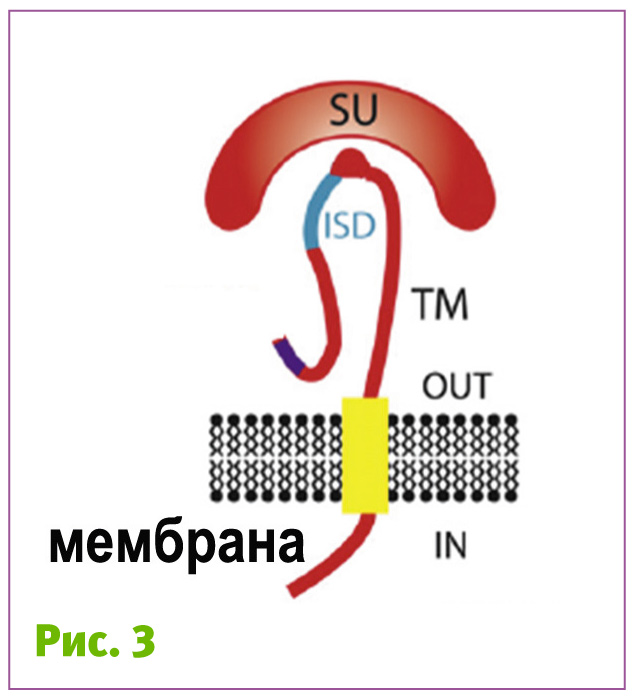
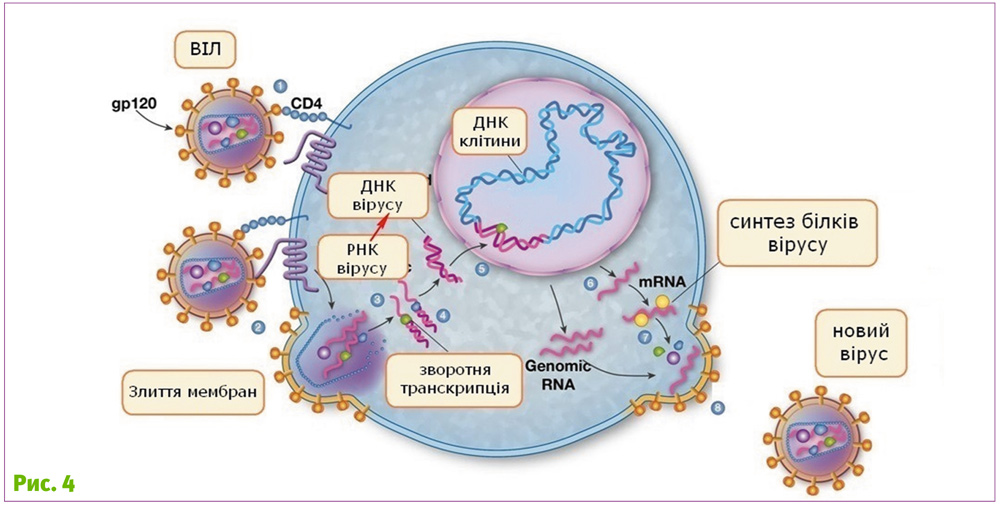
Вірусам, для того, щоб розмножитись, необхідно проникнути в інші організми. Для цього у кожного вірусу є свої пристосування. Наприклад, у ретровірусів, одного з багатьох сімейств вірусів, на оболонці є білки, що кодуються геном env (від англ. envelope – оболонка). Субодиниця SU (surface subunit, рис. 3) цих білків зв’язується зі специфічними рецепторами клітини і, завдяки цьому, вірус приєднуєтся до клітини.
Наступний етап – проникнення всередину клітини шляхом об’єднання оболонки вірусу та мембрани клітини. Для цього існує інша субодиниця – ТМ (transmembrane subunit). Вона одним кінцем постійно прикріплена до мембрани вірусу, а іншим, після того, як субодиниця SU зв'яжеться з рецептором клітини, занурюється в мембрану клітини. Потім ТМ білок вірусу «скручується», стягуючи мембрани вірусу та клітини ближче один до одного, що врешті призводить до їхнього злиття. А коли мембрани зливаються, вміст вірусу потрапляє у клітину.
Яскравий конкретний приклад – вірус імунодефіциту людини (ВІЛ, англ. HIV). Білок його оболонки має всі характерні складові: SU для нього – gp 120, а TM-білок – gp 41. Gp 120 взаємодіє з рецептором CD4 на поверхні деяких клітин (рис. 5). А gp 41 сприяє злиттю мембран вірусу.
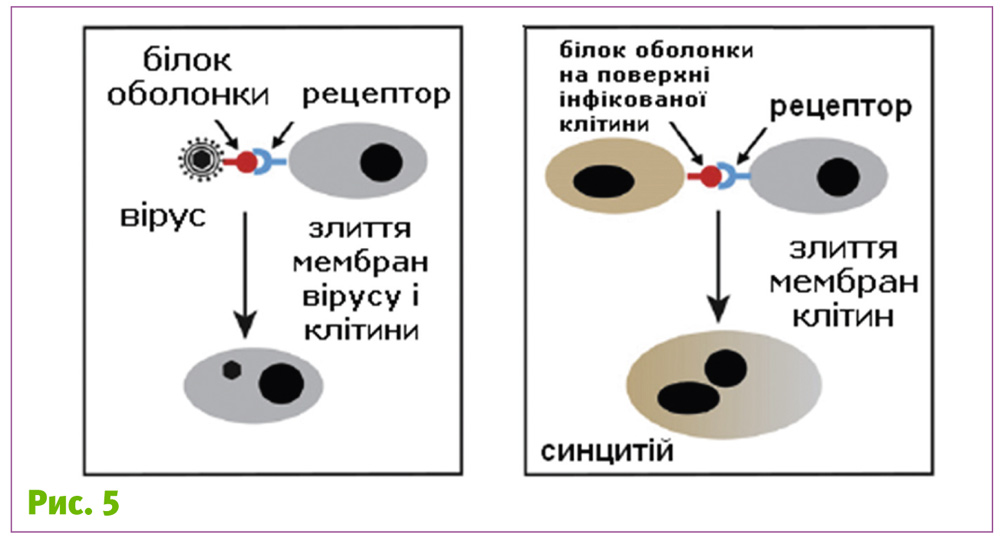
Як тільки вірус потрапив у клітину, він починає користуватися всіма її ресурсами. Але деякі з вірусів за допомогою того ж білка оболонки можуть об’єднувати ресурси декількох клітин! Вони примушують їх зливатись і утворювати синцитій! Той самий синцитій (від грец. syn – разом, і cytos – клітина) – багатоядерну структуру, утворену внаслідок злиття декількох клітин.
Це відбувається тому, що після об’єднання з вірусом на поверхні інфікованих клітин залишаються білки оболонки вірусу. Взаємодіючи з відповідними рецепторами на поверхні сусідніх клітин, вони сприяють їх злиттю, так начебто інфікована клітина – це великий вірус (рис. 5).
Особливістю ретровірусів є те, що вони мають здатність вбудовувати свою генетичну інформацію у геном хазяїна. Разом з власними генами клітини зчитуються також і гени вірусу, внаслідок чого синтезуються нові віруси, які здатні виходити за межі клітини та заражати нові. А вбудована вірусна ДНК може залишитися в клітині на все життя і при поділі клітини передаватися дочірнім.
Іноді буває так, що вбудовування ДНК вірусу в геном хазяїна відбудеться в клітині статевій, тоді вірусні гени будуть передаватися ще й від батьків до дітей. Такі «осілі» віруси, що містяться в геномі, називаються ендогенними ретровірусами – ЕРВ. Саме вони можуть «гастролювати», передаючись від предків до нащадків роками, і залишатися в геномі хазяїна, паразитуючи на ньому.
А тепер відповідь на одне з попередніх питань. У геномі людини знайдено саме такі декілька вірусних генів, які залишились нам у спадок від ретровірусів. Тепер ці гени кодують функціональні білки в плаценті. Саме їхні білки якраз відповідають за злиття клітин у синцитій. Мова йде про гени білків оболонок – гени env (Dupressoir et al. 2012).
Дивовижні речі! Вірусні білки, які допомагали вірусу, тепер працюють на наш організм. Важливо наголосити, що ці гени залишились активними. За декілька мільйонів років з моменту зараження вірусом, в його генах, що залишились поруч з нашими, зазвичай накопичується маса мутацій, що роблять вірус нездатним до розмноження та розповсюдження шляхом утворення нових вірусних часток. А у генів env, кодуючих білки плаценти, цих мутацій мало, тому вони й досі функціонують.
Отже, в плаценті є два білки, які кодуються вірусними генами env і відповідають за злиття клітин трофобласту. Штучне введення генів цих білків у інші клітини (там, де їх не буває), а внаслідок цього функціонування самих білків, викликало утворення синцитію. За що їх (тобто білки) так і прозвали – синцитин-1 та синцитин-2.
Так само як і всі вірусні env-білки, синцитини складаються з SU та TM частин і працюють вони по тому ж принципу, зв’язуючись з відповідними їм рецепторами на клітинах, сприяючи об’єднанню їх мембран.
Наскільки важливі синцитини? Невже наш організм не може впоратись без білків якихось вірусів? Якщо у ембріонів мишей «вимкнути» ген синцитину-А (аналог людського синцитину-1), тобто зробити його абсолютно неактивним, внаслідок чого перестане вироблятися функціональний білок, то такі ембріони помирають в утробі. Помирають через недорозвинення плаценти, в цьому випадку не відбувається формування шарів синцитію – структури, через яку здійснюється обмін речовин між матір’ю і плодом, структури, що знаходиться на межі двох світів.
Щоправда, коли «вимикали» ген синцитину-В (аналог людського синцитину-2), то спостерігали дещо іншу картину: клітини трофобласту, хоч і не зливались, але формували між собою дуже щільні контакти за рахунок інших білків – конексинів, – котрі компенсували таким чином недостачу синцитину та забезпечували тісну взаємодію між клітинами іншим способом. Завдяки цьому мишенята якось виживали, але почувались не так бадьоро, як здорові.
Як не крути, без вірусного спадку нам не обійтись. Звичайно, ці дивовижні білки не єдині із задіяних в утворенні та функціонуванні плаценти, є і багато інших дійових осіб (наприклад, білок ADAM) (Huppertz et al. 2006). Але судячи з усього, синцитини відіграють критичну роль.
Цікаво помітити, що об’єднання клітин трофобласту відбувається неперервно протягом всієї вагітності від початку імплантації. Ресурс синцитію необхідно постійно поновлювати, клітини нижнього шару трофобласту – цитотрофобласту – все більше і більше зливаються в синцитіотрофобласт. Це говорить про те, що роль багатоядерної структури дуже важлива. Більш того, через порушення процесів злиття клітин трофобласту у людини спостерігаються клінічні патології, такі як прееклампсія (Vargas A et al., Reprod Sci 2011).
Сам синцитій – це, грубо кажучи, багатоядерна величезна клітина. Головна його перевага – внаслідок відсутності клітинних мембран – швидкий транспорт речовин по всьому синцитію. Якщо він не утворюється (при вимкненні синцитину-В), клітини намагаються щільніше взаємодіяти між собою (підвищувати кількість звичайних білків взаємодії – конексинів). Що означає компенсацію і важливість саме властивості тісного контакту в синцитії. Але це ще не все.
Загадка трансплантації
Плацента походить від двох організмів, утворюючисьз клітин ембріона та клітин матері. Можна сказати, що це фізіологічна трансплантація, оскільки потім здійснюється функціонування генетично різних тканин фактично в одному органі. Але чому ж організм матері не відторгає ембріон як чужорідний об’єкт так само, як це відбувається при трансплантації? Чому не відторгається плацента з ембріоном протягом 280 днів у людини, а, наприклад, у слона навіть 660 днів? Адже для організму матері ембріон – це чужорідне тіло. Ще майже 40 років тому вже думали, а чи не в плаценті криється розгадка? Щоб це перевірити, здійснили експеримент за допомогою методу, який широко застосовується в трансплантології для підбору донора при пересадці органів, – культури змішаних лімфоцитів (КЗЛ). Метод КЗЛ використовують для перевірки сумісністі донора і реціпієнта, тобто приживеться трансплантант в організмі пацієнта, чи ні. Цю сумісність оцінюють по імунній реакції лімфоцитів пацієнта у відповідь на додавання лімфоцитів донора. І чим менше реагують клітини, тим краща сумісність. Відповідно, при повній несумісності лімфоцити пацієнта починають ділитись і проявляти імунну відповідь по відношенню до лімфоцитів донора. Так от, коли до таких несумісних клітин додавали матеріал, виділений з плаценти з білками мембран трофобласту, то поділу не спостерігалось. Білок трофобласту пригнічував імунну реакцію! (McIntyre & Faulk 1979).
Виявляється, справа все в тих же білках Env, запозичених від вірусів, і все в тих же властивостях, притаманних вірусам. Крім того, що віруси здатні викликати злиття клітин, об’єднуючи їх в один величезний ресурс, вони ще й пригнічують імунну відповідь в організмі, щоб не відбувалось відторгнення.
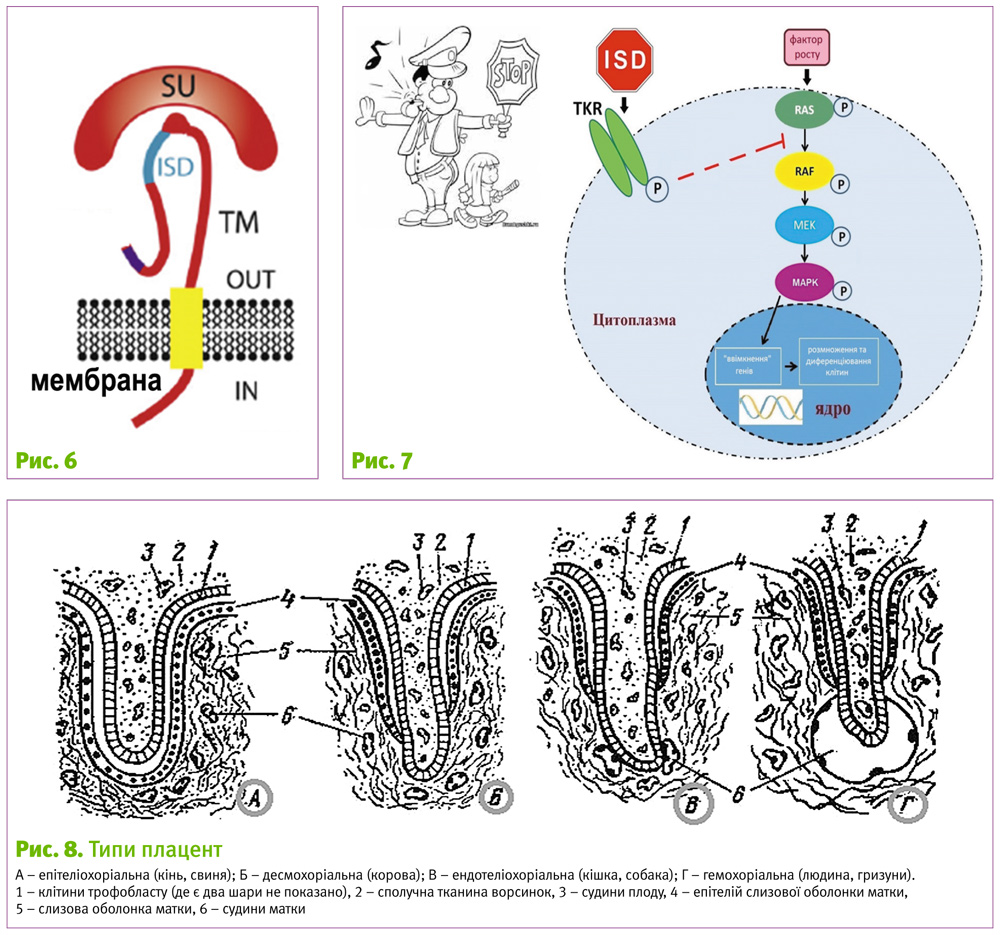
У білках Env для цього існують ділянки ISD (immunosuppressive domain, рис. 7).Передбачається, що ISD зв’язується з рецептором тирозинкінази – білка-регулятора двох сигнальних шляхів (зокрема, «Ras-Raf-MEK-MAPK», рис. 8).
Як наслідок, зменшується вироблення запального інтерлейкіну 12 – активатора клітин, що вбивають заражені і хворі клітини, а також підвищується синтез протизапального інтерлейкіну 10, пригнічується вироблення антитіл. Отже, загалом імунна відповідь пригнічується (Blinov et al. 2013).
На сьогодні така імунопригнічувальна властивість підтверджена для самих синцитинів. Введення гену синцитину пригнічувало відторгнення організмом штучно підсаджених пухлинних клітин, в той час коли пухлинні клітини без синцитину пригнічувались, не встигаючи розростись до пухлини (Mangeney et al. 2001).
Хоча, не варто стверджувати, що наявність синцитинів – єдина причина примирення тканин двох генетично різних організмів. У трофобласті майже немає молекул головного комплексу гістосумісності (МНС), а ті, що є, ховаються, або маскуються антитілами матері. Лише все разом – наявність імунопригнічувальних ділянок в синцитинах плаценти, мала кількість МНС і багато чого іншого – дозволяє організму матері спокійно відноситись до зародку, що прилаштувався в її організмі, не активуючи проти нього своєї оборони.
Критична роль білків вірусного походження Env доведена безсумнівно. Без цих білків не відбувається нормального розвитку плаценти, а з нею і плоду. Цікаве і відкрите питанням: а чи не вірусам ми завдячуємо взагалі появою плаценти як суспільного надбання? Певні факти цілком можуть свідчити на користь цієї гіпотези.
Наприклад, якщо у різних тварин прослідкувати за структурою плаценти на межі тканин материнського та ембріонального походження, то можна розгледіти деякий зв’язок. Чим більше виражено функціонування синцитинів, тим нахабніше себе веде трофобласт по відношенню до тканин матки, тим ближче він намагається потрапити до жаданого живильного середовища – крові (рис. 8, Carter 2012).
Здається, здатність клітин зливатись впливає на їх інвазивність. Здається, це синцитин робить так, що ембріон спокійненько прилаштовується в тканинах матері. Адже це ми зараз думаємо, що так і має бути, що дитині за всіма канонами належить жити у матері під крилом. А по суті? Інший, новий організм, для матері напівчужий, збирається користуватися її ресурсами – пити кров, що називається! До цього треба було вправно приноровитись.
Проте, є кілька «але», які піддають сумніву виникнення плаценти завдяки вірусам. По-перше. Якщо деякі гени env вимикати, плід помирає, ще не народившись. А плацента, хоч і дефектна, все одно формується! Значить, не лише гени env винні у розвитку плаценти. По-друге, існує ж і нормальна плацента без синцитію – епітеліохоріальна – і народжуються до сих пір нові свинки, такі рожевенькі і хороші, і нові конячки. Значить життя без синцитину існує.
Але є припущення, що плацентарність ссавців почалась зовсім не з цього «безсинцитійного» типу плаценти (рис. 8), де трофобласт тихо-мирно тільки влаштовується по сусідству з епітелієм матки, а вже з інвазивного типу – ендотеліохоріального чи навіть гемохоріального, – де трофобласт вбудовується в тканини. Останнє говорить про те, що можливо «безсинцитійна» плацента утворилась вторинно, коли функції набутих синцитинів були втрачені. Хоча, чи були гени env у найперших плацентарних – теж невідомо.
Ось така загадка. І все ж таки, як би там не було, на які б питання ми не відповіли, з упевненістю можна сказати одне. У нас і в організмах наших родичів-тварин (далеких чи близьких) є гени env, які містять інформацію про білки оболонки вірусів. Ці білки раніше слугували вірусам як інструмент для інфікування та пригнічення імунітету організму хазяїна. А тепер деякі з них служать нам для функціонування плаценти і називаються синцитинами. Вони забезпечують здійснення більш тісної та компромісної взаємодії між матір’ю та ембріоном.
І потрапили ці «просинцитинові» віруси до нас природним шляхом, інфікувавши наших предків та затишно влаштувавши власні гени поруч з нашими.
Наш розвиток, наша еволюція залежить від того, що нас оточує, від того, що з нами відбувається, в тому числі від вірусних інфекцій. Тому, якщо на ваш погляд сталось щось погане, не поспішайте засмучуватись, бо немає лиха без добра.
Перелік літератури знаходиться у редакції.
коментарів