

Відкриваючи скриньку Пандори
Імплантаційний парадокс ще в 1945 р. був описаний P. Viganо oднією фразою, що надалі стала афоризмом: «Бластоциста може віртуально імплантуватися у будь-якому місці людського організму, крім нерецептивного ендометрію». Дійсно, імплантація може відбутися у будь-якій тканині людського тіла, найчастіше без попередньої підготовки. У той же час, ендометрій відноситься до числа тих рідкісних тканин, де цей процес можливий лише за наявності так званого вікна імплантації, коли ендометрій максимально чутливий до рецепції ембріона, тобто коли він «дозволяє» зародку імплантуватися.
У тварин імплантація може бути відтермінована на невизначений час. Таким чином відбувається весняна синхронізація та відновлення поділу ембріону з одночасною диференціацією матки. Яскравою ілюстрацією є феномен «відтермінованої імплантації» у гризунів: бластоциста може кілька місяців перебувати в порожнині матки і чекати гормональних сигналів, що дозволяють імплантацію. В той самий час та ж бластоциста, поміщена в інше оточення (наприклад, в черевну порожнину), швидко активується та занурюється у навколишні тканини.
У людини вікно імплантації дуже мало вивчене з ряду причин, серед яких домінують технічні та етичні чинники. Насправді жодна модель in vitro не здатна відтворити складність клітинної взаємодії епітеліальних, стромальних та імунокомпетентних клітин, представлених в ендометрії протягом вікна імплантації. Практика біопсії ендометрію у людини в периімплантаційний період небажана, тому дані про імплантацію ембріона людини дуже обмежені. Доступна інформація заснована на роздільному вивченні ендометрію і бластоцисти на моделях in vitro.
Імплантація складається з двох феноменів: апозиція–адгезія, спільна для всіх ссавців, і пенетрація–інвазія, яку знаходять у гризунів та приматів. На першому етапі зародок розташовується у найзручнішому місці та розпочинає діалог з ендометрієм за допомогою цитокінів та іонів, а молекули адгезії інтегрини поверхні ендометрію утворюють зв’язки зі специфічними лігандами трофобласта, насамперед онкофібронектином, розпочинаючи «прилипання».
Адгезія трофобласта і ендометрію – справжній біологічний феномен. На рівні місця імплантації ці два типи епітелію різного генетичного походження (епітелій бластоцисти з одного боку і неадгезивна поверхня апікальної частини клітин епітелію з іншого боку) зливаються. Наступний крок – поступове занурення зародка у стромальний шар ендометрія, що закінчується до 10 доби від запліднення.
Для вивчення причин порушення процесів імплантації потрібно розуміти, що адекватна відповідь ендометрію на зустріч із зародком обумовлена насамперед якістю останнього. Ідеальною моделлю природного відбору є відсутність імплантації всіх зародків низької якості, насамперед анеуплоїдних. У присутності такого зародка включається прозапальна цитокінова відповідь, що блокує його адгезію та інвазію. За умови «здорового» зародка етапи імплантації можуть бути порушені не лише станами з боку ендометрію і матки, але й загальними розладами, у першу чергу, з боку систем – учасників діалогу «зародок–матка».
З точки зору варіантів взаємодії ендометрію з зародком доцільно використовувати термін «децидуальний фенотип». За умови адекватної селективності здорового ендометрію відбувається імплантація якісного зародка і подальший нормальний перебіг вагітності, тоді як анеуплоїдні ембріони втрачаються, насамперед, периімплантаційно. З точки зору кінцевого результату цікавим є термін «інапарентні вагітності» – тобто ті потенційні живонародження, які ніколи не стануть такими внаслідок природного відбору. Згідно даних JD. A. Delhanty, Frank Pellestor, 2011 з 100 ооцитів, що овулювали, народиться 31 дитина, решта 69% втратяться на різних етапах, але переважна більшість з них – периімплантаційно (мал. 1). 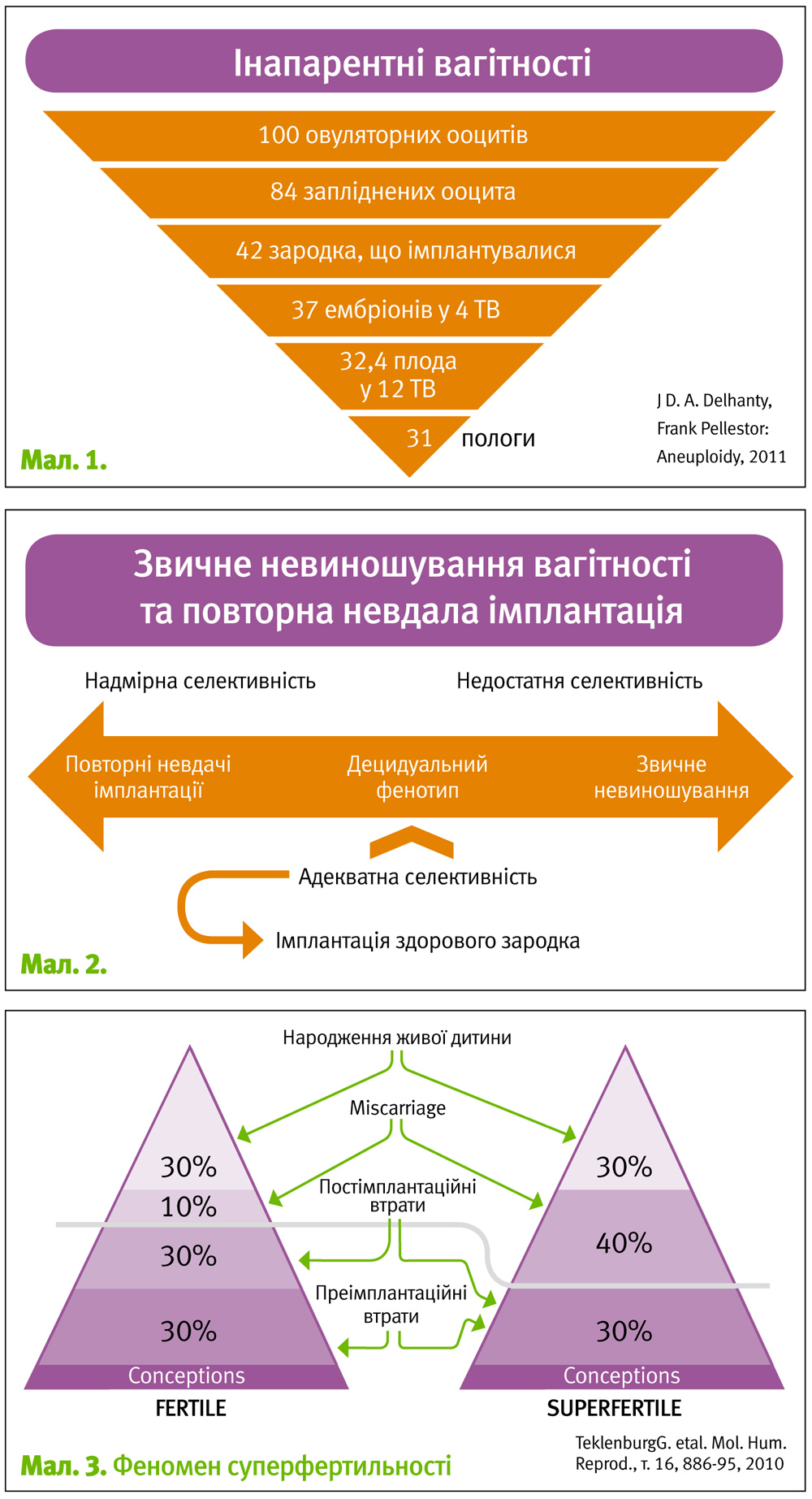
Близькі ж за причинами та проявами порушення репродукції у вигляді звичного невиношування та повторних імплантаційних втрат мають протилежні децидуальні фенотипові характеристики. Так, у першому випадку ендометрій відрізняється недостатньою селективністю («нерозбірливістю»), тому імплантуватися може значна кількість неякісних зародків, які у подальшому будуть втрачені як клінічні вагітності. Прогнозована ймовірність настання вагітності у таких пацієнток є більшою у порівнянні з нормою (так званий «феномен суперфертильності»). Протилежний децидуальний фенотип характеризується, навпаки, надмірною селективністю, в умовах якої навіть здорові зародки не можуть імплантуватися – клінічно це проявляється неплідністю (яка у даному випадку є відносною) та невдалими спробами ЕКЗ (мал. 2).
Кількість ембріонів, що втрачається таким чином, в обох випадках буде приблизно однаковою, проте втрачатимуться вони на різних етапах і матимуть різні клінічні прояви та, відповідно, назви.
Регуляція ендометріальної рецептивності
Рецептивність ендометрію визначається саморегульованим періодом часу, протягом якого епітелій ендометрію переходить у тимчасовий стероїдозалежний стан, що забезпечує адгезію бластоцисти на його поверхні. У людини епітелій набуває цей статус одночасно з децидуалізацією строми. Цей процес обумовлений, насамперед, дією прогестерона після відповідного підготування 17-в естрадіолом. Це є вікно імплантації, що триває з 4–5 до 9–10 дня після підвищення рівня прогестерона (19–24 дц).
Яєчникові стероїди відіграють роль медіаторів у діалозі «ембріон–матка» і модулюють експресію чисельних факторів росту, цитокінів та молекул адгезії у материнської-плодовому просторі. Прогестерон забезпечує секреторну активність ендометрія, забезпечуючи сприятливе середовище для розвитку ембріона та його імплантації. Секреторна трансформація ендометрія, викликана прогестероном, супроводжується каскадом експресії генів, що полегшують або, навпаки, обмежують імплантацію ембріона.
Поява зрілих піноподій збігається з утворенням плато концентрації прогестерону в крові. У жінок з нормальним менструальним циклом піноподії з'являються між 19–21-м днями менструального циклу, під час передбачуваного вікна імплантації. Кількість естрогенових і прогестеронових рецепторів змінюється протягом менструального циклу. Існує значна різниця між кількістю рецепторів у периовуляторний і периімплантаційний періоди – від піку в епітелії та стромі на початку секреторної фази до майже відсутності їх в епітелії на 20–22 день циклу на тлі максимальної концентрації прогестерону та естрадіолу в крові. Вважається, що саме різке зниження кількості стероїдних рецепторів в епітелії призводить до появи на їхній поверхні білків адгезії (інтегринів), а також підвищує чутливість до факторів росту.
Фундаментальну роль в імплантації відіграють також хоріонічний гонадотропін людини (ХГл), інгібін, активін, релаксин, кальцитонін. Можливо, ХГл – одна з перших складних молекул, синтезованих ембріоном. Він бере участь на всіх етапах діалогу «ембріон–матка». Лептин і його рецептори продукуються ендометрієм і плацентою. Він модулює інвазивні характеристики цитотрофобласту.
Останнім часом увагу багатьох вчених прикуто до вивчення ролі цитокінів у системі репродукції людини. Фактор, що інгібує лейкемію – один з найцікавіших цитокінів з точки зору репродуктивної функції тварин і людини.
ФІЛ, або leukemia inhibitory factor (LIF), продукується різними видами клітин, включаючи фібробласти, клітини гемопоетичного ряду, гепатоцити, ендотеліальні клітини, сенсорні і автономні нервові клітини, остеобласти, астроцити, деякі необластичні клітини. Потенційними індукторами синтезу ФІЛ є інтерлейкін-1 (IL-1), фактор некрозу пухлини (TNF), епідермальний фактор росту (EGF), трансформуючий фактор росту (TGF). Було виявлено, що ФІЛ in vitro підтримує ембріональні стовбурові клітини у недиференційованому стані, стимулюючи їхню диференціацію, але не впливаючи на проліферацію клітин.
У самок мишей з інактивованим ФІЛ-геном процес фертилізація яйцеклітин не порушений, однак останні виявилися нездатними до імплантації.
ФІЛ присутній практично в усіх клітинних структурах і тканинах репродуктивної системи людини, включаючи яєчники, маткові труби, матку, плаценту. Рівень ФІЛ в ендометрії низький у проліферативну фазу, підвищується після овуляції і залишається відносно високим майже до кінця менструального циклу, знижуючись з початком менструації до базального рівня. У складних взаєминах між ембріоном і материнським організмом ФІЛ відіграє важливу роль як медіатора, так і стимулятора.
Цитокін бере участь у процесах трансформації трофобласта у різні його форми: в інвазивний цитотрофобласт або високодиференційований синцитіотрофобласт, він здатний модулювати рівень хоріонічного гормону, стимулюючи або пригнічуючи його продукцію у трофобласті залежно від тривалості, дози або часу впливу.
ФІЛ здатний індукувати продукцію протеїназ та фібронектина клітинами трофобласта, приймаючи таким чином участь у регуляції інвазії трофобласта.
Інший важливий для імплантації цитокін – гранулоцитарно-макрофагальний колонійстимулюючий фактор росту (ГМ-КСФ), що сприяє росту лейкоцитів та розвитку трофобласта за рахунок індукції, проліферації та диференціювання стовбурових клітин.
В експерименті у мишей з нуль-мутацією ГМ-КСФ порушуються процеси імплантації та плацентації, вдвічі збільшується частота вад розвитку та втрат вагітностей, вдев’ятеро збільшується частота СЗРП, кількість нащадків внаслідок пре- та постнатальних втрат зменшується на 25%, при цьому плоди чоловічої статі значно чутливіші до відсутності фактору росту. У жінок із ЗНВ спостерігається значне зменшення концентрації ГМ-КСФ у сироватці крові (Perricin et al., Am. J Reprod. Immunol 2003), а також експресії мРНК ГМ-КСФ в ендометрії (Jasper et al., 2007).
ГМ-КСФ знаходиться у достатній кількості в клітинах фолікула, ендометрії та епітелії фалопієвих труб, а зародок експресує сам цитокін та рецептори до нього. У період овуляції кількість ГМ-КСФ регулюється естрогенами, а потрапляння сіменної рідини у репродуктивний тракт викликає різке збільшення ГМ-КСФ та прозапальних цитокінів ендометрієм. Другий пік експресії цитокіна спостерігається у період імплантації, після чого він секретується у клітинах трофобласта, децидуальній тканині, плаценті. Г-КСФ сприяє збільшенню клітинної маси та зниженню рівня апоптозу. У нормі ендометріальний апоптоз негативно корелює з концентрацією естрадіолу в сироватці крові, а саме достовірне збільшення інтенсивності апоптозу спостерігається при падінні концентрації естрадіолу і прогестерону в пізній лютеїновій фазі. Ембріон є активним учасником процесу імплантації: наявність бластоцисти в стадії позиціонування оберігає ендометрій від апоптозу. Доведено, що навіть ембріони, які не досягли стадії бластоцисти, знижують кількість апоптотичних клітин у культурі ендометріальних епітеліальних клітин. Однак далі, на стадії адгезії, бластоциста індукує паракринну апоптотичну реакцію. Найімовірніше, система запуску апоптозу Fas / Fas-L задіяна у подоланні бар'єру для імплантації бластоцисти.
Імунологічна регуляція імплантації
Поряд з вищевикладеними механізмами, в імплантації ембріона задіяні імунологічні процеси.
Імунологічні зміни під час вагітності відбуваються на загальному та місцевому рівнях. Згідно останніх досліджень, для нормального перебігу вагітності необхідний баланс між, з одного боку, Treg із зсувом в бік Т2 та, з іншого боку, зменшенням впливу Т1 та Т17. Вирішальне значення для нормальної імплантації та перебігу вагітності мають ПК-клітини, які поділяють за фенотипом на два класи за експресією маркера CD56: CD56dim та CD56bright. CD56dim є первинною ланкою реагування на чужородні антигени, CD56bright відрізняються високою експресією маркера та продукують регуляторні цитокіни. Для нормального перебігу вагітності необхідний зсув в сторону ПК2 та зменшення цитотоксичності ПК1. Проте достеменно невідомо, чи дійсно впливає кількість та цитотоксичність ПК периферійної крові на перебіг вагітності, оскільки основні імунологічні зміни під час вагітності відбуваються на місцевому рівні. Існує певна відмінність періферійних ПК (пПК) від маткових ПК (мПК). мПК складають 40% всіх лейкоцитів ендометрію у проліферативну фазу, та до 70 – у секреторну фазу та на початку вагітності. Регуляція чисельності мПК відбувається прямим (гормональним) шляхом через рецептори до стероїдів та непрямим шляхом, що опосередкований цитокінами. мПК виробляють чинники локальної імуносупресії, глікоделін та галектин-1, а також фактори росту (VEGF, MIP-1β), що викликає перебудову судинного русла децидуальної оболонки, та цитокіни, що полегшують інвазію трофобласта. мПК контактують з позаворсинчатим трофобластом та експресують рецептори до його антигенів. Позаворсинчатий трофобласт занурюється у децидуальну тканину, забезпечуючи фізичне прикріплення та адекватний пацентарний кровоплин.
Доведено підвищення концентрації мПК лютеінової фази в жінок, що мали 3 та більше невдалі спроби ЕКЗ при переносі якісних ембріонів або ≥3 втрат вагітності (King et al., 2010). Зниження кількості ПК також є несприятливим для репродуктивної функції, оскільки призводить до порушення ремоделювання спіральних артерій та недостатньої антигенної презентації ембріона з наступним порушенням імплантації. Проте, незважаючи на наведені раніше дані, суперечливим залишається питання визначення кількості ПК периферичної крові. Це пов’язано із наявністю циркадних ритмів, та суттєвої різниці при визначенні частки або кількості ПК. За даними мета-аналізу 22 досліджень Seshadri et al., 2013, точність клінічної діагностики на підставі такого визначення сумнівна.
Гемостаз та імплантація
Вплив на імплантацію тромбофілії різного походження обумовлений надмірною активацією факторів згортання на тлі пригнічення фібрінолізу. Наслідком такого дисбалансу в згортальній та протизгортальній системах та ендотеліопатії є локальні геморагії та мікротромби у ділянці інвазії бластоцисти, недостатня глибина інвазії трофобласта та неповноцінна плацентація.
Проте погляди щодо впливу природжених тромбофілій на імплантацію є суперечливими. Значна частина досліджень демонструє вищу, ніж у популяції, ймовірність імплантації та високу частота пологів у носіїв мутації фактору V Leiden та протромбіну (Rudick B. Fertil. Steril., 2009).
Більше згоди щодо ролі антифосфоліпідних антитіл у генезі порушень репродукції. Кардіоліпінові і фосфатидилсеринові антитіла специфічно викликають пошкодження трофобласта, порушують формування синцитіуму, змінюють продукцію людського хоріонічного гонадотропіну (ХГл), порушують інвазію трофобласта. АФА підвищують експресію інгібітора активатора плазміногену 1 (PAI-1) і тканинного фактора (TF), що посилює протромботичні механізми і знижує активність фібринолізу, що також призводить до дефектів імплантації та зниження глибини децидуальної інвазії трофобласта, пошкодження мембрани ендотелію, порушення балансу в ланці природних антикоагулянтів та інгібіторів згортання і провокування тромбофілічного стану.
Дослідження маркерів рецептивності
Останнім часом проводяться активні дослідження з пошуку маркерів рецептивності ендометрію. Існує велика кількість публікацій, присвячених прогностичному значенню ультразвукового моніторингу, доплерографії, тестів функціональної діагностики, визначення рівня сироваткового прогестерону (неінвазивні методи). До малоінвазивних методів належить визначення концентрації різних речовин у змивах з порожнини матки.
Інвазивні методи полягають у дослідженні біоптату ендометрію, отриманого за допомогою гістероскопіі або пайпель-біопсії із застосуванням морфологічних методів, імуногістохімічних методів, скануючої електронної мікроскопії, геноміки, протеоміки та метаболоміки.
Найбільш доступним і широко застосовуваним є морфологічне дослідження ендометрію: у жінок зі звичними неефективними імплантаціями найчастіше виявляються гіперплазія та гіпоплазія ендометрія, ендометрит та дисхроноз.
Ще у 1950 р. Noyes R. була запропонована морфологічна хронологічна система характеристики ендометрію, що дозволяє виявити невідповідність між його передбачуваною хронологічною та реальною гістологічною структурою. Незважаючи на численні дискусії, хронологічне датування – один з найбільш доступних і ефективних методів оцінки функціонального стану ендометрію, а точність діагностики можна підвищити, використовуючи строгі критерії хронологічних параметрів менструального циклу, при цьому для зменшення суб’єктивності оцінки рекомендоване незалежне оцінювання принаймні двома морфологами.
Наступний етап – імуногістохімічні дослідження експресії ядерних стероїдних рецепторів. Відомо, що для повноцінної децидуалізації важлива не стільки абсолютна кількість стероїдних гормонів, скільки наявність певної кількості функціонально повноцінних рецепторів ендометрію. Знання рецепторного статусу ендометрію дозволяє прогнозувати його відповідь на замісну гормональну терапію. З метою імуногістохімічного дослідження експресії ядерних стероїдних рецепторів використовують E-tegrity-test (Beta-3 integrin), який дуже ефективний, проте фінансово затратний. Науковий інтерес становлять маркери проліферації та апоптозу: Bcl-2, p53, індекс апоптозу (ІА), експресія маткового LIF, CD56 +, CD16-, CD3 + (маткові натуральні кілери), маркер хронічного ендометриту CD138 +, а також маркери децидуалізаціі: пролактин та IGFBP1.
Методом скануючої електронної мікроскопії (СЕМ) поверхневого епітелію ендометрію виявлено наявність характерних випинань апікальної частини плазматичної мембрани у період вікна імплантації, які отримали назву піноподіі, а їх визначення було запропоноване у якості одного з маркерів сприйнятливості ендометрію.
Генетичні дослідження ендометрію в період «вікна імплантації» принесли багато несподіваних результатів. Виявилося, що залученість генів у циклічну трансформацію ендометрію величезна. Показано, що при відкритті «вікна імплантації» в ендометрії посилюється експресія близько 395 генів, функція яких пов’язана з з клітинним метаболізмом, залозистою секрецією, диференціюванням клітин, міжклітинною комунікацією та природженою імунною відповіддю. Одночасно знижується експресія близько 186 генів. На підставі отриманих даних було запропоновано оцінювати рецептивність ендометрію у конкретної пацієнтки за допомогою вивчення експресії генів у передбачуваний період імплантації. Метод ERA (Endometrial Receptivity array) використовує дослідження на основі генетичного чіпу експресії 238 генів, що відповідають за ендометріальну рецептивність.
Вибір тактики ведення пацієнток з підозрою на ендометріальний чинник порушення імплантації полягатиме у детальному аналізі наявних даних та, безперечно, критичній оцінці можливостей. Інформація про стан ендометрію у пацієнток зі звичним невиношуванням та невдалими спробами ЕКЗ, проаналізованого морфологічно та імуногістохімічно, може бути вкрай важливою для призначення відповідного лікування, від протизапальної терапії до зрошення ГМ-КСФ або проведення гістерорезектоскопії.
Дані про зміну імунологічного статусу, наявність тромбофілічних станів, можуть бути використані для додаткового лікування, що не завжди є ефективним з точки зору доказової медицини. Окремі важкі випадки пошкодження ендометрію та порушення його рецептивності можуть бути, на жаль, вирішені лише за допомогою замінного материнства.
![]()
коментарів